

环境变化威胁着全球的生物多样性,尽管一些迁徙动物已经通过调整迁徙路线和时间做出反应,但这些反应通常不足以跟上近期环境变化的步伐。在这些变化中,气候变化正在改变迁徙的最佳时间,导致迁徙动物与其食物来源之间的物候不匹配。另一方面,栖息地恶化威胁着连接繁殖地和非繁殖地的网络的完整性,尤其是当它导致栖息地大部分或完全丧失时。即使只占用很短的时间,迁徙网络中的特定停歇点也可能对维持整体迁徙活动和整个种群至关重要。因此,在关键停歇点,即使是低水平栖息地恶化也可能导致快速的种群下降甚至灭绝,特别是当迁徙动物缺乏足够的适应能力来应对这些变化时。因此,预测迁徙动物适应环境变化的能力是生态学中的一大关键挑战。
过去六十年来,东亚-澳大利西亚迁徙路线由于污染、过度开发和土地复垦,经历了严重的栖息地退化和丧失。数百万只鸻鹬科鸟类沿着这条迁徙路线每年迁徙,依赖沿海的潮间带泥滩进行休息和补充能量。在黄海地区,栖息地退化最为严重,估计潮间带丧失超过50%。然而,这个特定地区在迁徙期间承载了该迁徙路线上的大部分鸻鹬科鸟类。许多鸻鹬科鸟类种群的迅速下降很可能是这些变化的结果。
迁徙鸟类,尤其是鸻鹬科鸟类,在世界上大多数主要迁徙路线上都在下降。然而,即使在东亚-澳大利西亚迁徙路线上,下降的幅度也存在很大差异:一些物种的年下降率高达11%(例如,弯嘴滨鹬,Calidris ferruginea),而其他物种似乎相对稳定(例如,红颈滨鹬,Calidris ruficollis)。虽然下降的确切原因尚不清楚,但可能是由不同地点的环境变化速度和幅度与每个物种的固有迁徙策略相互作用驱动的。迁徙策略至少部分依赖于体型,包括鸟类在一次飞行中能飞多远、能积累多少体能储备以及飞行的能量成本。迁徙策略主要在于使用的中途停留地点数量和它们之间的距离。环境变化可能对具有不同迁徙策略的物种产生不同影响。例如,如果一个具有单一停歇点和长途迁徙鸟类在其唯一的停歇点面临栖息地退化,它要么必须改变迁徙策略,要么将面临灭绝,而如果一个停留地点是一个连接良好的多停歇地点网络的一部分,那么失去一个停留地点可能只需要对整体迁徙策略进行轻微调整。
这项研究开发了一个建模框架来预测鸻鹬科鸟类的最佳迁徙策略,关注生态和物种特定特征,包括体型和生理能力,并将这些特征与东亚-澳大利西亚迁徙路径上的环境条件变化联系起来。研究对象为五种体型对比鲜明的鸻鹬科鸟类(Scolopacidae),共预测了历史(1960年代)、当前(2010年代)和未来(2060年代)的最佳迁徙策略,并将模型预测与2013年至2019年间收集的追踪数据(n=61)进行了比较。共测试了以下假设:a)体型较小的物种需要沿迁徙路径分布更多的中途停留地点,从而可以缓冲特定地点的不利变化,b)当前体型较小物种的迁徙更接近其预测的最佳迁徙,而体型较大的物种则需要在迁徙中进行更多重大改变,并且在最近的快速环境变化期间更有可能表现出次优迁徙。另外确定了迁徙路径上各个停留区域的相对重要性,并预测在未来环境变化下,这些区域的重要性可能会转移到新的地区,并需要建立新的停留地点。
模拟最佳迁徙
这项研究开发了一个状态依赖的动态迁徙模型,以计算迁徙鸟类从非繁殖地到繁殖地北迁时最大化其适应性的迁徙决策,其中特别关注春季迁徙,因为它与个体适应性有明确相关性。在每个时间段中,鸟类可以选择留在当前地点觅食或迁徙到下一个地点,取决于身体储备、时间以及当前和后续地点的预期条件。该模型针对五种密切相关的鸻鹬科鸟类的特定种群进行了参数化(下图),这些鸟类在东亚-澳大利西亚迁徙路径上有相应追踪数据(n=61):斑尾塍鹬(Limosa lapponica baueri)、大滨鹬(Calidris tenuirostris)、红腹滨鹬(Calidris canutus,5只rogersi和1只piersmai)、弯嘴滨鹬和红颈滨鹬。在研究中,计算了最佳决策,即适应性最大化,其中包括在获取能量和避免捕食之间的权衡,并权衡靠近繁殖地的好处(或成本)与留在当前地点的好处(或成本)。
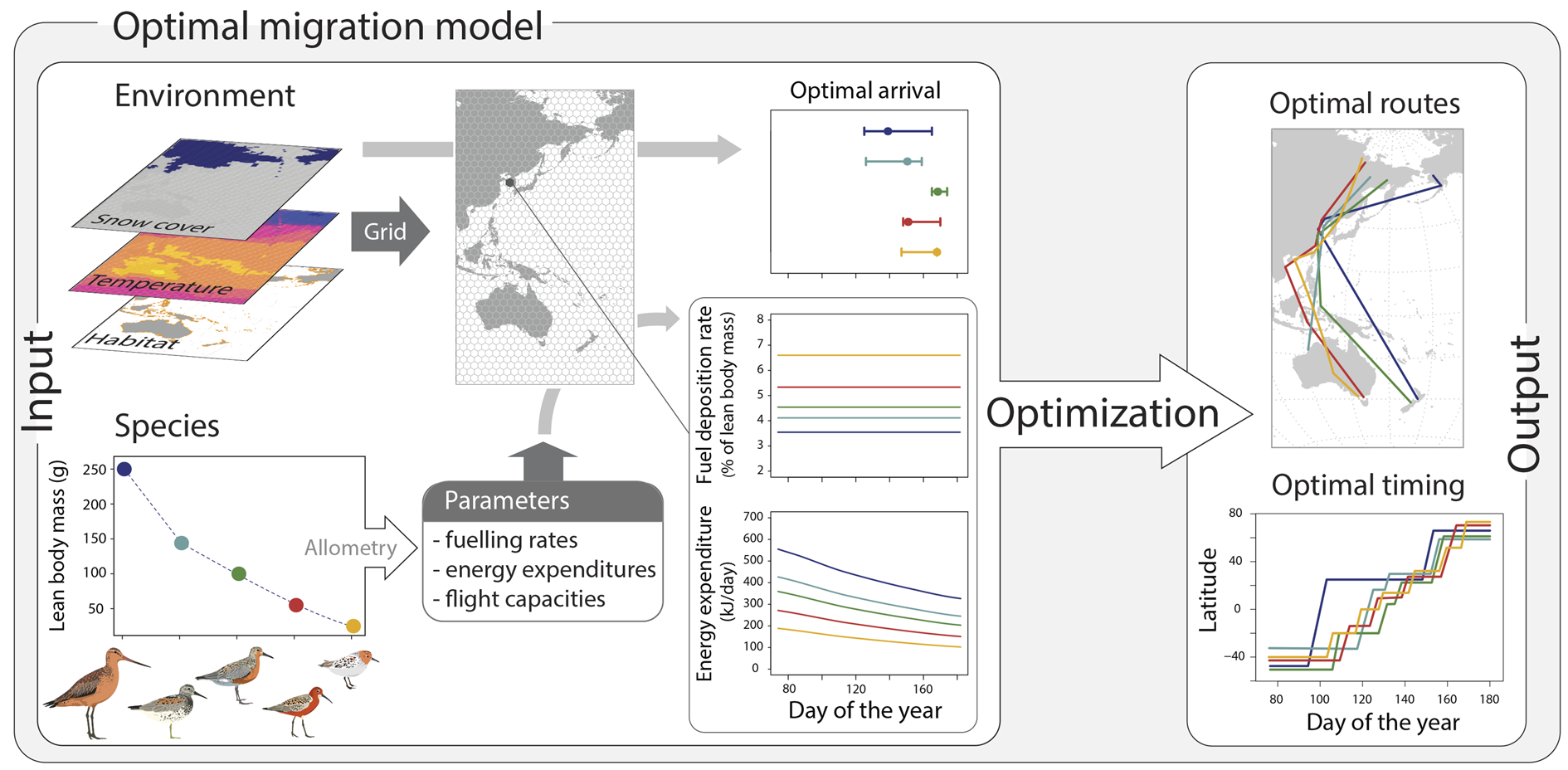
最相关的模型参数定义了物种特定的生理特征,例如能量维持(温度依赖的能量消耗)、最大能量补充率和飞行成本。东亚-澳大利西亚迁徙路径被分为一个六边形网格(直径400公里,面积138,564平方公里)。每个单元的环境条件分别定义为过去(1960年代)、当前(2010年代)和预测的未来条件(2060年代),在分析中使用了三组变量——每日温度、潮间带泥滩范围和融雪日期——假设i)环境温度影响能量消耗,ii)潮间带泥滩范围表征食物可用性和觅食选择,iii)融雪日期决定繁殖地的最佳到达时间,它们的具体值来自于过去和现在条件的遥感和气候的分析结果,以及对未来的预测。
基于迁徙追踪轨迹及其特定的主要非繁殖地和繁殖地,研究为每个时期(1960年代、2010年代和2060年代)模拟了200条物种特定的最佳迁徙路线。从所有模拟轨迹以及迁徙追踪结果中,提取了表征迁徙策略的数据:迁徙持续时间(从春季出发到到达繁殖地的时间)和迁徙距离(从主要非繁殖地到繁殖地的距离),同时基于模型的空间网络对中途停歇地点数量进行分组,以便对模型预测和迁徙追踪路线进行比较,每个地点的停留时间也用于推导个体和物种特定的迁徙策略。
追踪迁徙结果
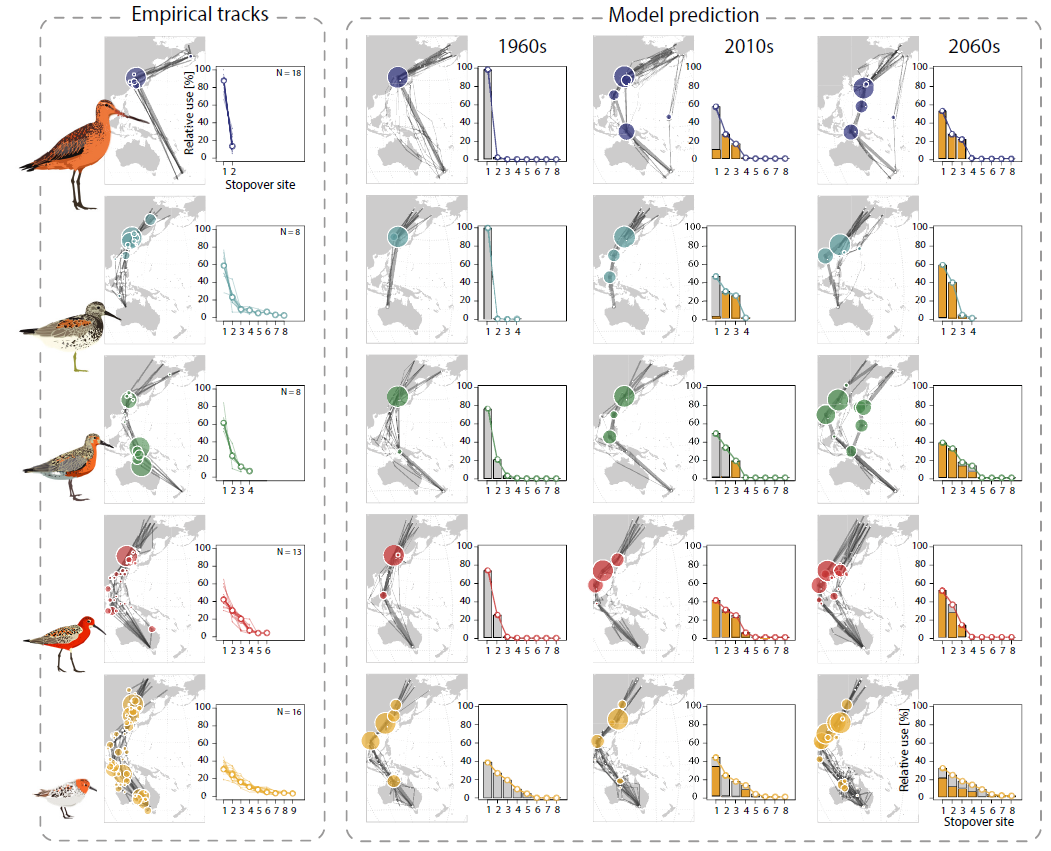
迁徙追踪数据显示了不同物种特定的春季迁徙策略(下图左),包括距离、停留地点数量以及每个地点的相对停留时间。斑尾塍鹬作为体型最大的物种,(体重250克),迁徙的距离最长(16,200 ± 550公里),采用了极端的“jump”策略,仅在黄海的一个主要停歇点停留,个体在整个迁徙过程中平均花费了88.2%的时间。只有很少的一部分时间(11%)花在靠近主要停留地点的第二个地点。
大滨鹬(体重144克)和红腹滨鹬(体重105克)使用了多个停歇点(4至8个地点),其中一两个地点的停留时间相对于其他地点更长。大滨鹬的主要停留地点位于黄海地区,占总迁徙时间的78.8%(55.4%至94.5%),红腹滨鹬广泛使用了北澳大利亚(例如卡奔塔利亚湾)和印尼南部的海岸线,占总迁徙时间的61.3%(52.6%至81.9%)。
两种体型较小的物种,弯嘴滨鹬(体重55克)和红颈滨鹬(体重23克),通常使用更多的停留地点(分别为3至6个和4至10个),且分布范围更广,并倾向于在这些地点上更加均匀分配时间(下图)。
模型预测比较
该模型预测了这些物种的不同迁徙策略,这些策略通常也会随着时间的推移而发生显著变化,但红颈滨鹬除外(上图最后一行)。将经验轨迹与模型预测进行比较,发现对于两个较小的物种,迁徙追踪路线当前(2010 年代)的预测更接近,而对于三个较大的物种,与过去(1960 年代)的预测结果更接近(上图 )。通过比较三个不同时间段的模型预测和经验轨迹在停留地点停留时间的分布,证实了这一点(下图A),对于三种体型较大的物种,在过去预测的相似度较高,对于弯嘴滨鹬,当前和未来的相似度较高,而对于红颈滨鹬,过去和现在预测的相似度没有差异。
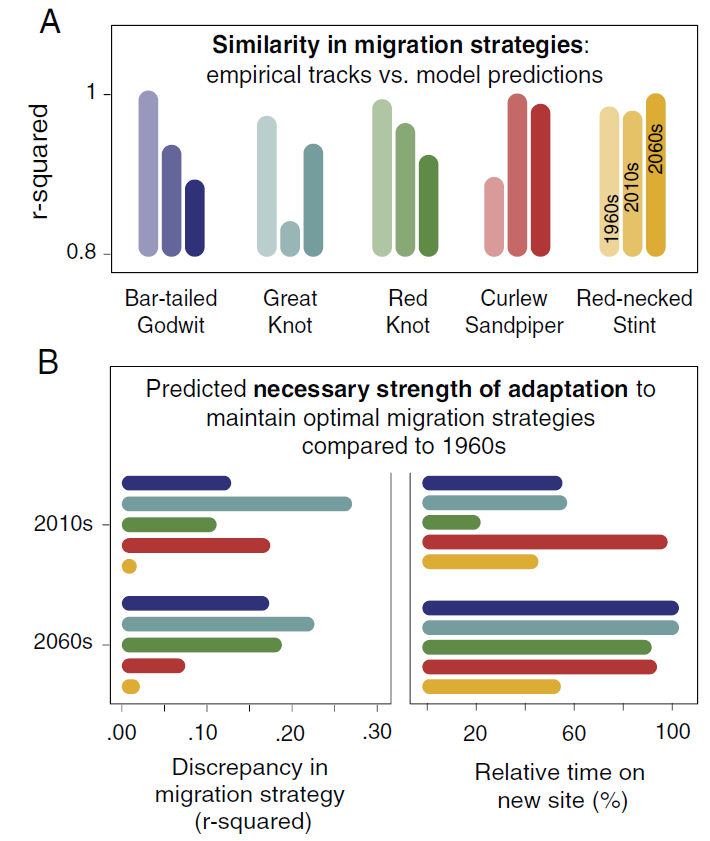
通过将当前(2010 年代)和未来(2060 年代)情景的预测与过去(1960 年代)情景的预测进行比较,量化了维持最佳迁徙策略所需的适应性(上图B左)。大滨鹬需要最高的适应性才能保持在最佳范围内。弯嘴滨鹬和斑尾塍鹬也需要较高的适应性,其次是红腹滨鹬,而红颈滨鹬不需要对其迁徙策略进行重大改变。在过去条件和未来情景的比较中,可以发现所需的适应性与体型有很强的相关性,这表明,适应的需要对于三种体型较大的物种来说是最大的,而对于弯嘴滨鹬和红颈滨鹬来说则要小得多。
除了红颈滨鹬外,所有物种的预测迁徙持续时间都随着时间的推移而增加(下图),过去与未来预测的相对增加:斑尾塍鹬 159%、大滨鹬 185%、红腹滨鹬 170%、弯嘴滨鹬 119%、红颈滨鹬 93%)。在弯嘴滨鹬中,预测的迁徙持续时间在目前条件下最长(为过去最佳迁徙的 165%)。预测的迁徙持续时间更长是因为个体离开越冬地的时间明显提前(过去和现在情景之间最明显的差异),同时也略早到达繁殖地(由于气候变化导致北纬地区的最佳到达窗口提前)。
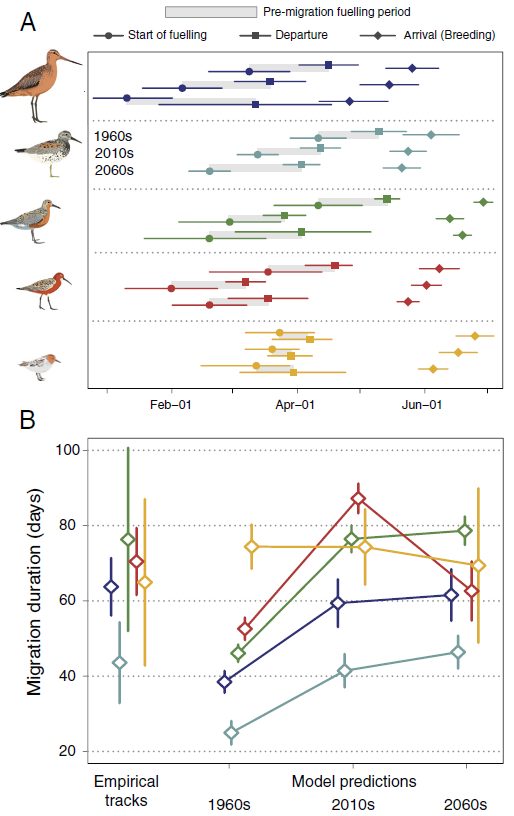
三个模型预测中,在不同纬度停留的相对时间发生了变化。在过去的条件下(20 世纪 60 年代),大多数个体依赖黄海地区,尤其是渤海(~37°N 至 41°N)。这种对黄海的依赖在现在(2010 年代)已经发生了变化,在未来(2060 年代)的条件下,这种依赖将更加明显,届时鸟类将越来越多地使用更靠南(20°N 至 33°N 和赤道附近)的替代地点,并且分布更加分散。
这项研究通过将模型预测与迁徙追踪路线进行比较,展示了在气候变化条件下,迁徙所需的改变,以及在物种之间的差异。总体而言,体型较大的物种需要更根本的改变以保持最佳策略,例如使用完全不同的地点和路线,而较小的物种不需要过多的调整。这个框架提供了一个强大的工具来识别由于多种环境变化而导致的迁移行为所需的适应。
参考文献
[1] Lisovski S, Hoye BJ, Conklin JR, Battley PF, Fuller RA, Gosbell KB, Klaassen M, Benjamin Lee C, Murray NJ, Bauer S. 2024. Predicting resilience of migratory birds to environmental change.[J] Proc Natl Acad Sci U S A.121(19):e2311146121.

