

为了深入理解植物群落的多样性是如何维持的,我们首先需要了解限制性资源的竞争机制。在生态学领域,当限制性资源变得稀缺时,竞争者的生长速度通常会减缓,而资源的枯竭更会缩短竞争者的生长时间——本研究将这一机制称为“时间竞争(competition for time)”。值得注意的是,现代群落生态学的理论基础和我们对物种共存问题的理解框架,都建立在竞争会降低生长率的隐含假设之上。然而,最新的理论研究显示,在众多自然群落中,“时间竞争”可能是最主要的竞争方式。这一发现意义重大,因为在物种竞争时间的情况下,多样性的维持会自然达到一种权衡。
本研究首先通过一个简单的物种相互作用模型,从概念上引入了时间竞争的概念。接下来,本研究在地中海地区的一年生草地上开展了一项实验,旨在验证时间竞争是否确实是野外生态系统中一个重要的竞争机制。实验结果显示,物种在竞争加剧时,更倾向于通过缩短其寿命来应对,而非降低其生长速度。综上所述,时间竞争作为一种维持生物多样性的机制,可能一直被忽视了。
研究背景
了解塑造植物群落结构的驱动因素,对于预测这些系统如何响应人为干扰、气候变化及物种入侵具有至关重要的作用。光、氮、水等资源已被证实会限制植物生物量的产生,因此,普遍认为对这些资源的争夺是塑造植物群落的主要过程之一。这也解释了为什么评价植物群落结构时,最具影响力的理论都聚焦于资源竞争,并对优势地位、物种共存、群落演替及物种入侵等重要问题做出了关键预测。
尽管资源竞争的核心地位鲜有质疑,但对这一过程的认知方式,可能会深刻影响其在塑造群落结构及应对全球变化方面的作用。特别是,对限制性资源的竞争会降低个体的生物量增长速度,同时竞争也会减少可用于生物量增长的时间——这种机制定义为“时间竞争(competition for time)”(图1)。当生物在连续的竞争中相互削减生长时间时,时间竞争就会对种群动态产生长远的影响(图2)。例如,这些竞争过程可能表现为不耐阴的林木在受到干扰后对光照的争夺,或者是一年生植物在冬季雨季后对水分的竞争。
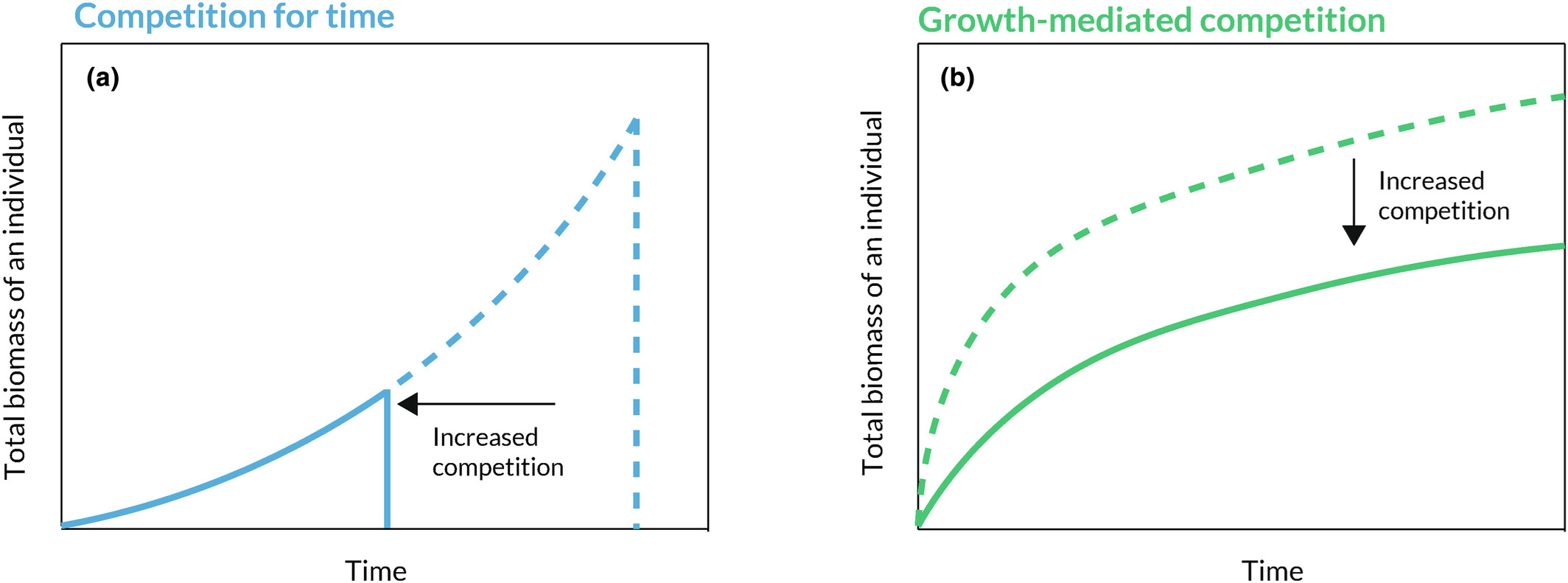
图1: 在时间竞争和以生长为媒介的竞争条件下个体生长轨迹的比较。两个面板都显示了竞争加剧对个体累积生物量的影响。(a)说明了在时间竞争条件下,竞争者密度的增加如何导致个体提前完成生长,从而在不影响生物量增长速度的情况下减少总生物量的积累。(b)说明了当竞争通过降低生长速度来表现时,个体在所有时间点的生长速度都会减慢,因此生物量总量会减少,而个体的寿命不会改变。
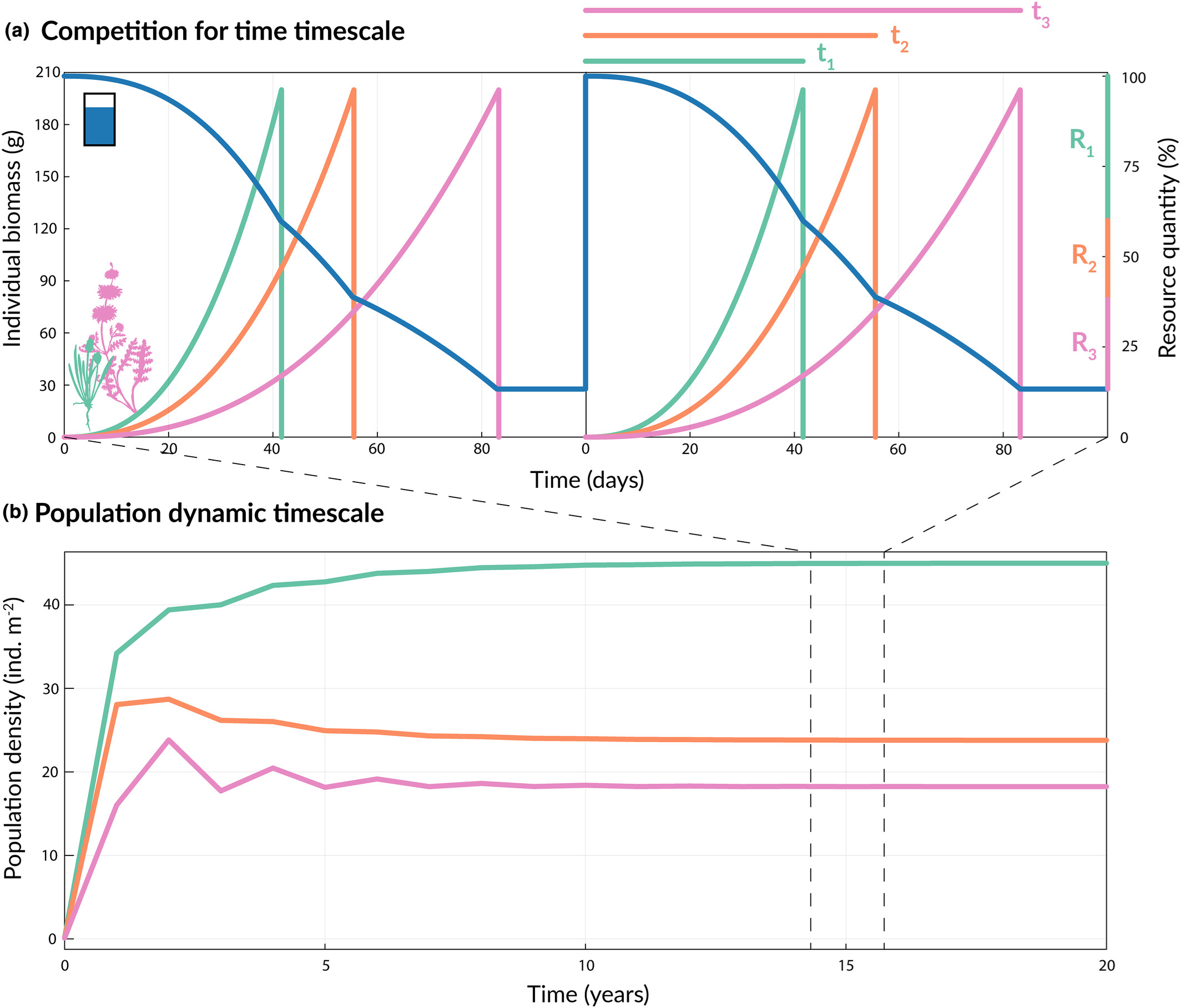
图2: 理论结果表明时间竞争是如何在竞争过程中发生的(a),但由于竞争的重复发生,时间竞争最终会在更长的时间尺度上影响种群动态(b)。
由于在这些系统和其他生态系统中竞争的反复性,时间竞争在更长的种群动态时间尺度上可能导致物种排斥或共存。例如,在一年生植物系统中,若动态竞争持续多年,可能会导致某些物种最终被排斥出群落。同样,降低生物量增长率的竞争也会影响多年的种群动态。然而,仅关注较短的种群动态时间尺度(如在一年生植物系统中的数年)的模型,必然会忽略对竞争的短期反应及其对自然界植物动态的影响。事实上,如Lotka-Volterra、Beverton-Holt和MacArthur消费者资源模型等经典群落动力学模型,都将竞争简单地视为降低种群增长的因素。
在常见的以生长为媒介的竞争模型中,能在限制性资源的最低水平上生长的物种占据主导地位,这导致共存的内生机会非常有限。在这种情况下,共存被认为依赖于某种外部机制来破坏竞争排斥,例如对多种限制性资源的特化、依赖密度的天敌攻击或时间储存效应。在过去的半个世纪里,群落生态学的一个重要研究焦点就是识别这些机制、它们在自然界中的出现位置以及它们对全球变化下生物多样性的影响。
然而,当竞争减少了可用于生长的时间时,维持多样性的权衡就会自然地产生。这是因为物种有机会将时间划分为不同的时段,在这些时段中每个物种都是最强的竞争者。这种划分通常是由物种在获取或容忍有限资源方面的差异造成的。当物种在生长和寿命之间进行权衡时,时间的划分相对于种间竞争而言尤其能加强种内竞争。这种权衡往往源于生态生理对生长和资源利用的限制。在这种权衡下随着寿命较短、生长速度较快的物种的退出,时间被划分为多样性递减的时期,从而为速度较慢的竞争者提供了时间上的庇护所。如果这一过程能够定期重置例如通过干扰来实现,那么高多样性就能长期保持下去。物种竞争时间时出现的权衡可能是自然界物种共存的一个被忽视的机制。
物种可能在各种生态系统中争夺时间,且这种竞争的范围正在不断扩大
时间竞争的一个典型例证是光照有限的森林中的次生演替。在这一过程中,资源需求量较大的先驱物种在受到干扰后会逐渐被高大但生长缓慢的物种所取代。这类系统中会出现时间竞争,因为随着高大个体的密度增加,矮小个体会更早地被淘汰,进而影响其整体的繁殖能力。尽管生长与寿命之间的权衡在维持演替森林高度多样性方面的作用已为人熟知长达一个多世纪,但这种机制在次生林演替之外的场景中很少被提及。然而,最近的理论研究显示,时间竞争可能在更广泛的生态系统中发生,包括那些没有演替动态的系统。
最近在模拟水分限制条件下植物生长的生态生理学研究中取得了进展,这些研究指出水分受限的植物群落也可能存在时间竞争。具体来说,这些群落展现出一种类似于生长季内演替的特征。在此过程中,植物并非因被森林中更高大的个体压制而死亡,而是在降水间歇期,为应对干燥的土壤条件而关闭气孔、停止生长。时间竞争之所以产生,是因为竞争者消耗了共享的水资源,导致个体比在无竞争环境下更早地停止生长。在这样的系统中,物种对干燥土壤条件的耐受性差异引发了一种连续的生长停止模式。当物种竞争时间时,促进共存的生长-寿命权衡机制与其他时间共存机制在几个重要方面存在差异。其中最主要的区别在于时间结构的来源。其他显著的时间共存机制,如储存效应,依赖于物种对波动的温度、降水、营养供应或其他外部因素的特定反应,以产生物种间的时间偏移。然而,在物种竞争时间的情况下,环境的变化几乎完全是内生的。例如,在森林中,光照的波动受高大竞争者密度变化的影响;在水分有限的植物群落中,气孔关闭的时间(以及植物生长的持续时间)由竞争者的蒸腾率决定。如果在生长和寿命之间进行权衡,这种内生的时间结构将有利于稳定的共存。因此,与其他明确涉及时间的共存机制相比,时间竞争在性质上更类似于相对非线性的关系。
时间竞争基于三个核心要求:(1) 物种能够从活跃的生长状态转变为非活跃状态;(2) 这种转变的时间是竞争的函数;(3) 处于非活跃状态的物种不会对活跃状态的物种构成竞争威胁。只要满足上述三个条件,即便转变是渐进的,时间竞争依然会发生,但这会引入以生长为媒介的竞争要素。
光照有限的次生演替案例,阐释了这些要求如何在竞争者之间实现时间的分配。受到扰动后,所有物种的个体会在大致相同的时间从种子开始生长。随后,当物种被更高大的竞争者超越时,它们便会死亡,仅留下休眠的种子或耐荫的新个体,而这些新个体对树冠层树木不产生影响。由于树种相对于竞争者的高度差异,树种的寿命也不尽相同,并且由于所有树种最初都是共存的,林分的生命周期被划分为多样性逐渐下降的阶段。
为了阐释密度依赖性的功能形式如何有利于这种机制的共存,本研究考虑了最简单的时间竞争模型:即离散世代的植物群落争夺单一的限制性资源。尽管这个模型仅描述了自然界中可能出现时间竞争的群落的一个子集,但其启示意义却具有广泛的适用性;而且该模型只需稍加修改,便可适用于更为复杂的系统。这个简单模型表明,在寿命较短的物种具有较高生物量增长率的情况下,共存的可能性会大大增加(图3a)。
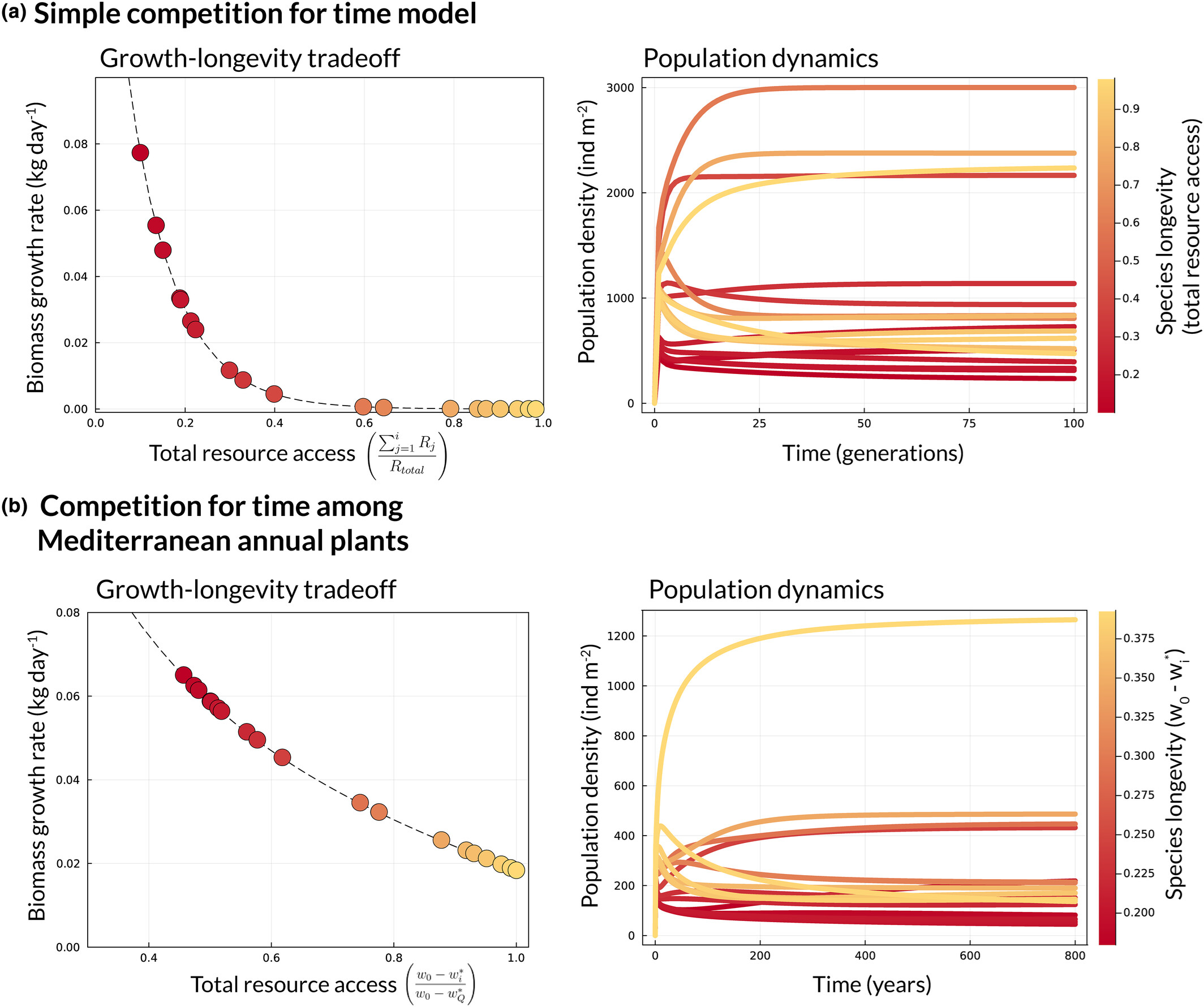
图3: 时间竞争简单模型中,生长-寿命权衡是如何保持高度多样性的。
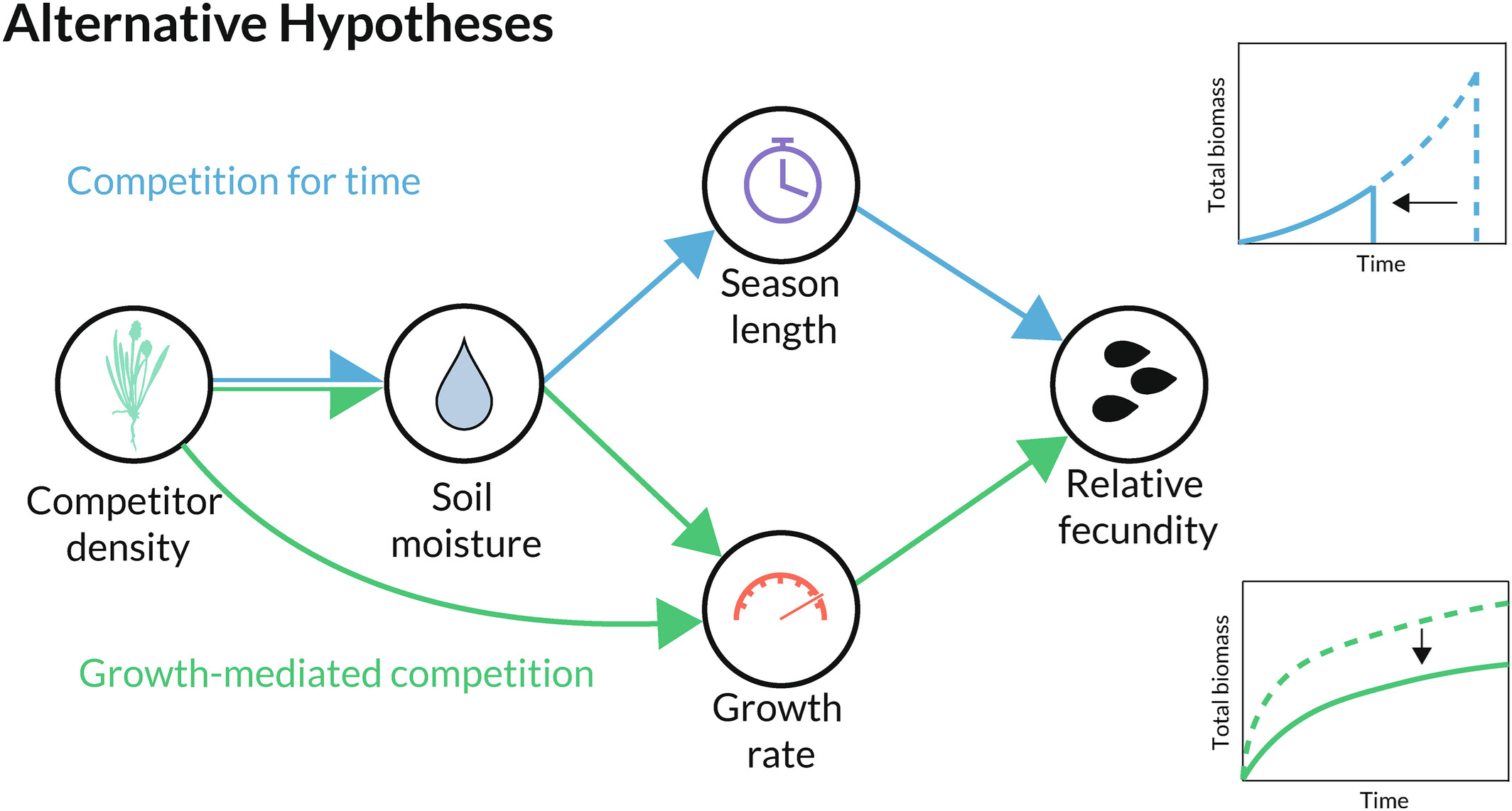
图4: 时间竞争实验中测试的替代假设。蓝色路径(上图)描述了地中海一年生植物竞争时间时的因果路径:竞争者密度增加导致土壤水分供应量降低,进而使个体在生长季节提前衰老,最终降低其繁殖力。绿色路径(底部)描述了当这些物种仅通过降低生物量生长速度进行竞争时的预期因果路径:由于土壤水分减少或其他原因,竞争者密度增加会导致个体生物量积累更慢,从而损害其总繁殖力。
物种竞争时间的实验证据
在非演替系统中对时间竞争进行实证测试,对于验证时间竞争的广泛存在性具有重要意义。为此,本研究开展了一项野外实验,旨在评估地中海一年生植物(其共存通常与演替不相关)中是否存在时间竞争。具体来说,探究了以下两个问题:(1)当物种在竞争水分时,水是否会限制个体的生物量增长率或其增长的持续时间?(2)物种是否遵循共存所必需的生长与寿命之间的权衡?
为了解答这些问题,本研究将五种加州一年生植物的目标个体分别种植在两类地块中:(a)单独种植特定竞争物种的地块,或(b)未种植竞争物种(作为对照)的地块。在一个生长周期内,对每个目标个体的生长状况、停止生长的时间点以及最终的种子产量进行了详细记录。
研究结果表明,竞争主要表现为生长季节长度的缩短,而非生长速率的降低(图5-6)。在五个重点物种中,有四个物种的竞争者密度对寿命的估计效应为负值(图5)。相比之下,仅有一个物种的生物量增长率因竞争而下降(图5)。

图5: 比较竞争者密度对生长季长度的影响(时间竞争,左图)和竞争者密度对生物量增长率的影响(以生长为媒介的竞争,右图)的实证分析结果。
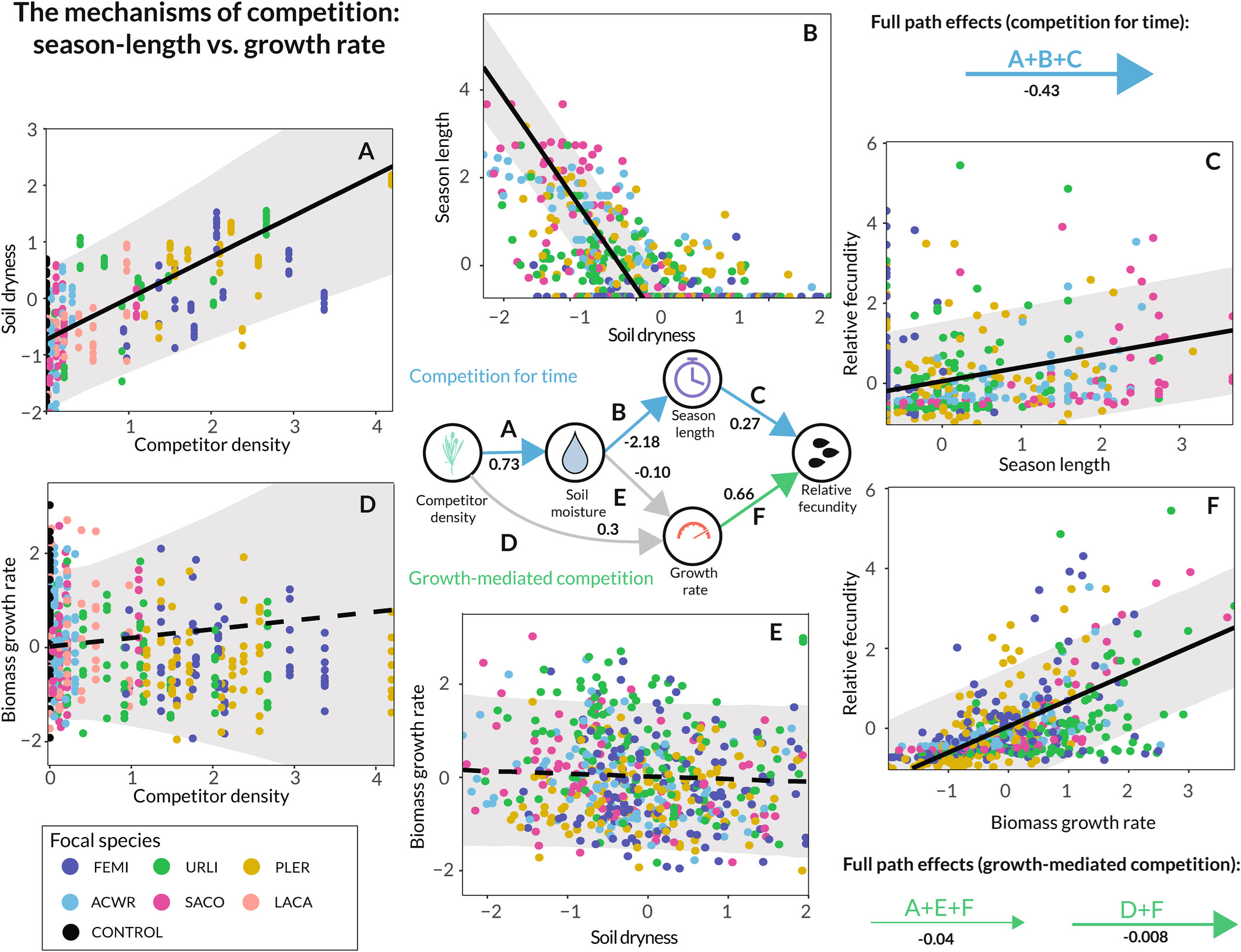
图6: 贝叶斯路径分析结果。
水分竞争对重点个体繁殖力的影响,主要是通过缩短物种的生长季长度(图6a-c),而非降低生长速率(图6d-f)来实现的。竞争者密度的增加与土壤条件更为干燥有关,而更干燥的土壤条件又与更短的季节长度有关,进而导致繁殖力降低(图6)。
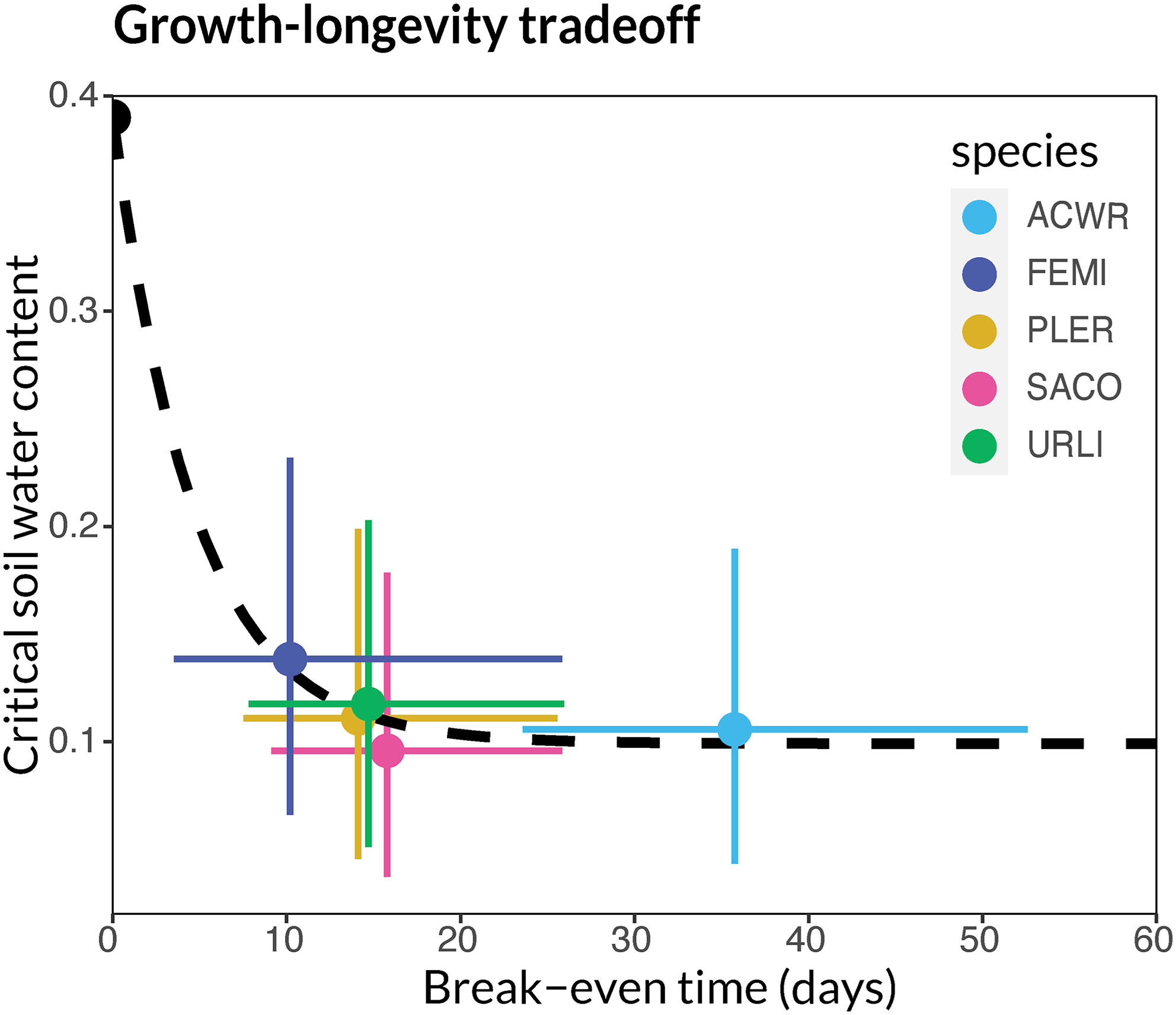
图7: 在田间试验中观察到的不同物种在盈亏平衡时间和临界土壤含水量之间的权衡。
本研究创新性地引入了时间竞争作为植物群落内的一种重要竞争机制,并详细阐述了当物种在生长与寿命间进行权衡时,时间竞争是如何有助于共存的。此外,本研究还通过野外实证研究,证实了时间竞争在植物生长、繁殖及物候结构中的关键作用。这一发现表明,时间竞争在维持多样性方面的作用可能远比之前认知的更为广泛。在实证研究中,所有物种在面临竞争者消耗土壤水时,相较于无竞争对手的个体,其生长会提前终止,从而导致生物量积累减少,种子产量也随之降低。
这一发现暗示了时间竞争作为一种竞争机制可能具有更广泛的重要性。未来的研究应致力于确定其确切的普遍程度。如果这种机制和相关的生长-寿命权衡被证实是普遍存在的,那么物种共存或许就不再是一个生态悖论了。

